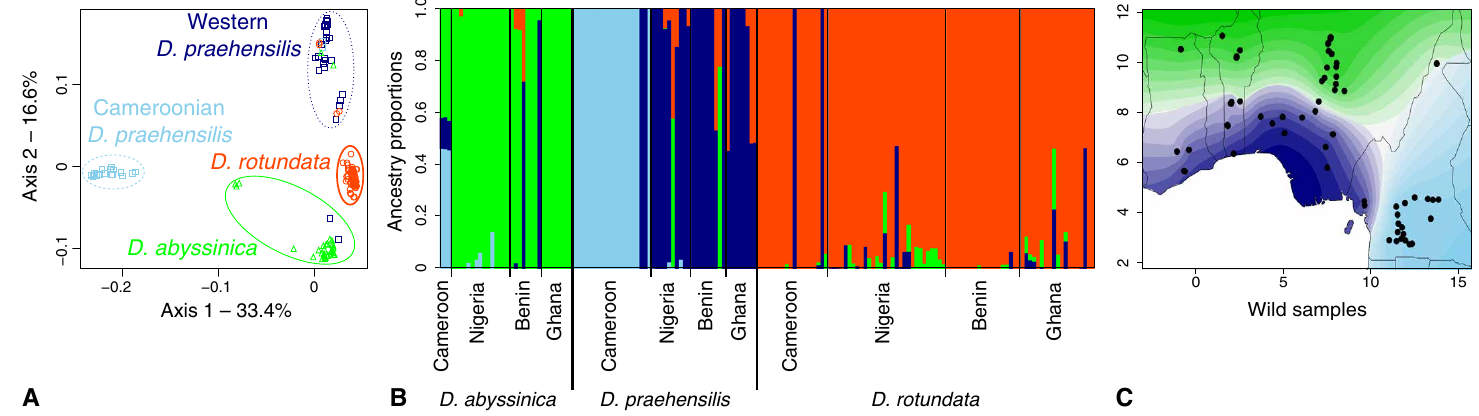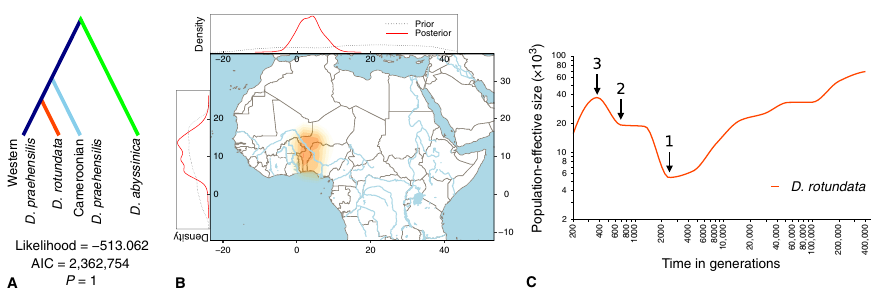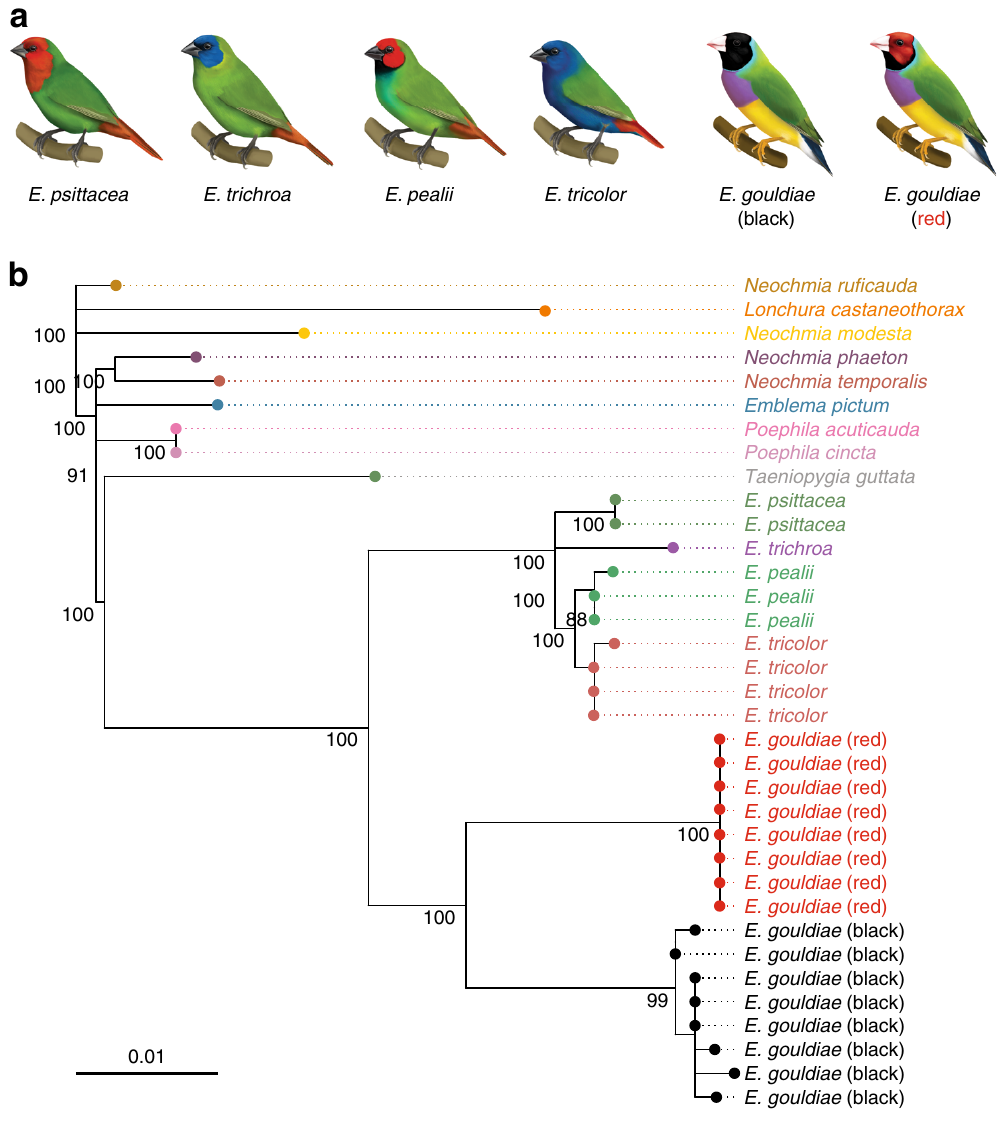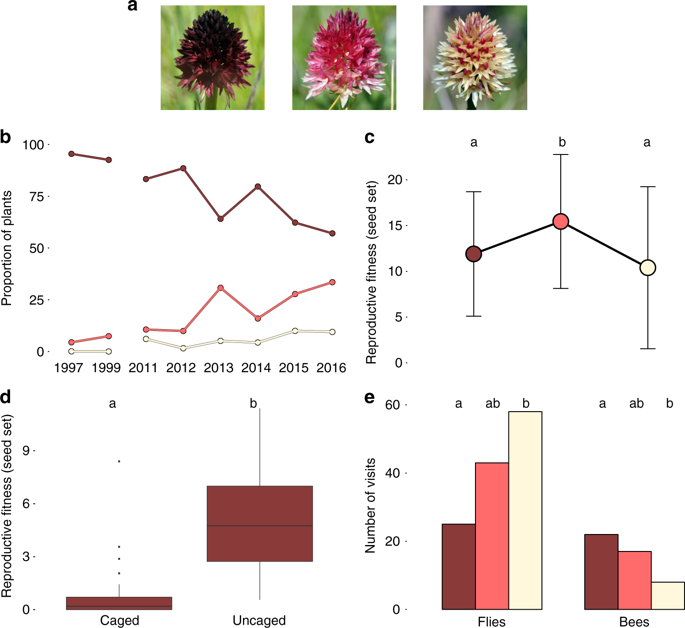
Le réchauffement climatique représente un défi majeur pour les populations naturelles de plantes. Les modèles théoriques de génétique des populations prédisent que les populations végétales répondront par une sélection positive sur des mutations pré-existantes initialement en fréquence faible avant que l’accumulation de mutations délétères n’entraîne l’extinction de la population. Ces prédictions théoriques n’ont que très rarement eu l’occasion d’être vérifiées.

Amidonnier sauvage (Triticum dicoccoides) (Source : Roger Culos, Wikipedia, publié sous licence CC BY-SA 3.0)
Le collectif Israélo-Canadien formé de Yong-Bi Fu et de ses collaborateurs a cherché à comprendre les mécanismes génétiques de réponse au réchauffement climatique chez des populations sauvages. Ce travail est décrit dans un article en Open Access publié dans la revue Proceedings of the National Academy of Sciences of the United States of America (PNAS) en septembre 2019. Dix populations d’amidonnier sauvage (Triticum dicoccoides Koern.), une forme de blé primitif, ont été échantillonnées en Israël, à la fois en 1980 puis 28 ans plus tard, en 2008, période sur laquelle les températures ont augmenté et les précipitations ont diminué en Israël. L’exome, c’est-à-dire les parties du génome codant pour des protéines, a été comparé entre ces populations. Les changements ont été étudiés en terme de mutation, sélection, diversité et différentiation entre populations.
Par rapport au premier échantillonnage en 1980, les populations de 2008 dans leur ensemble subissaient davantage de sélection, leur diversité génétique avaient diminué et elles portaient un fardeau génétique accru. Ce fardeau génétique représente la proportion de mutations non synonymes, c’est-à-dire qui changent la séquence en acides aminés, souvent de manière délétère. Toutefois, les résultats variaient selon les populations étudiées et certaines avaient même réussi à sélectionner des mutations bénéfiques, déjà présentes en fréquence faible en 1980 ou survenues après. La comparaison de populations poussant dans des conditions environnementales contrastées aussi bien que la fonction des gènes ayant le plus divergé au cours de la période d’étude indiquent que, sur les populations étudiées, l’augmentation de la température aurait eu un impact plus important que la baisse des précipitations.
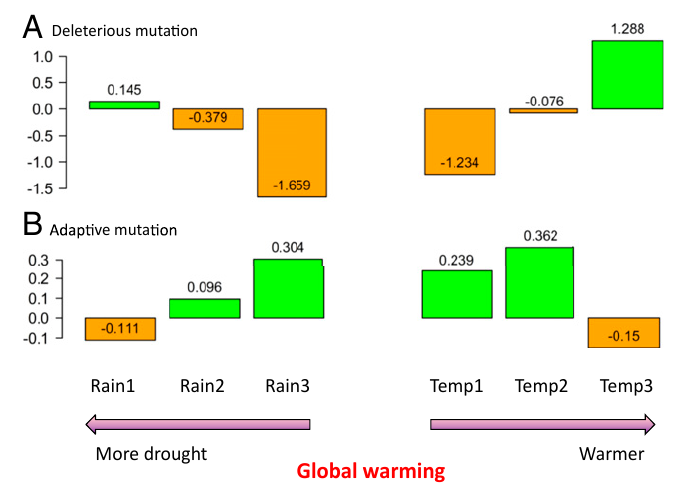
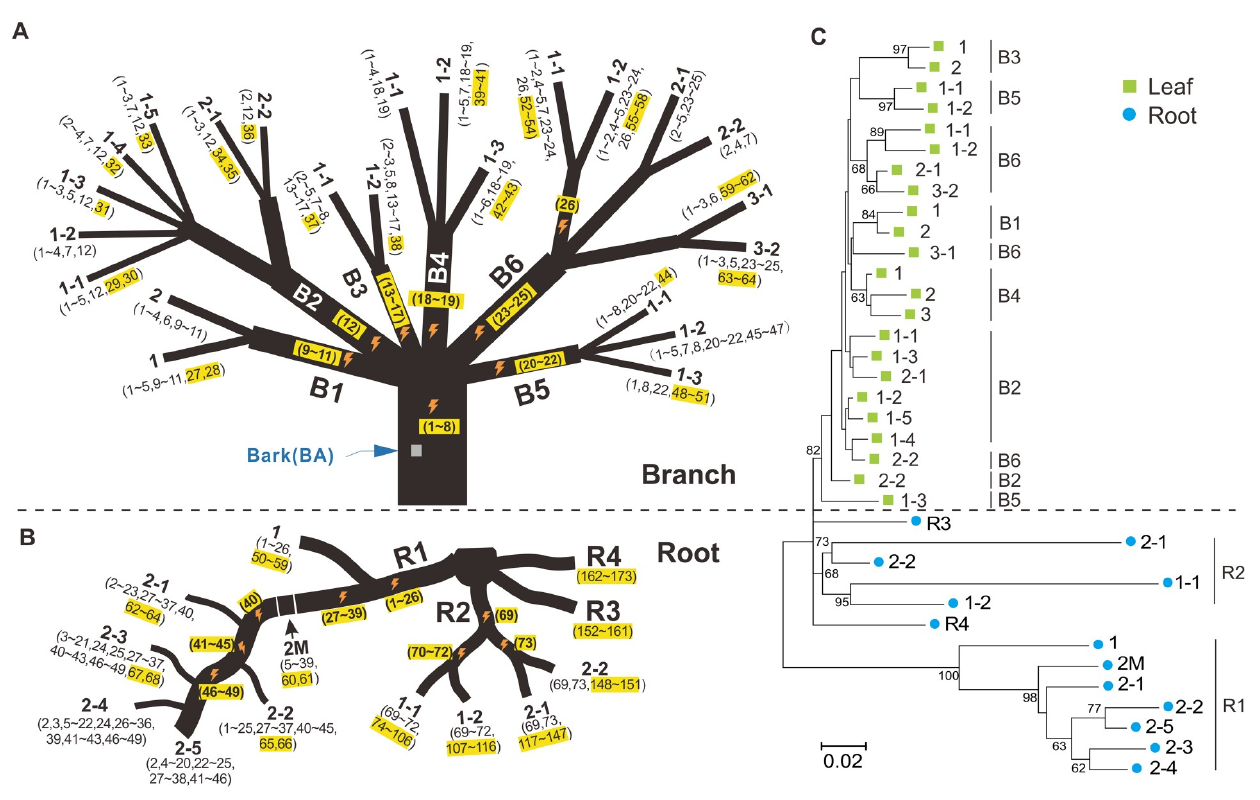
Effet du réchauffement climatique sur les mutations des populations d’amidonnier sauvage. Sur ce schéma, les populations ont été regroupées en six groupes selon la quantité de précipitations annuelle de la zone d’échantillonnage (gauche) ou de sa température moyenne annuelle (droite). Les populations des zones les plus sèches et celles des zones les plus chaudes sont celles qui avaient accumulé le plus de mutations délétères (A; fardeau génétique) et le moins de mutations avantageuses (B), qui leur auraient permis de s’adapter.
Si cette étude donne des indications sur le potentiel adaptatif des populations étudiées à résister au réchauffement climatiques en cours ou au contraire leur vulnérabilité, elle apporte surtout et plus généralement des espoirs quant à de futurs travaux de modélisation plus réalistes pour comprendre les facteurs d’adaptabilité ou de vulnérabilité des populations naturelles au changement climatique. De tels modèles pourraient être utilisés comme outils d’aide à la décision pour la prise de mesures de préservation des populations naturelles.
Ces populations naturelles ne sont pas que des « mauvaises herbes ». Ainsi, du fait de sa proximité avec le blé, l’amidonnier sauvage représente une ressource potentiellement utile pour l’amélioration génétique du blé concernant les stress abiotiques (tolérance à la sécheresse, chaleur, etc.) ou biotiques (tolérance aux pathogènes et ravageurs). Toutefois, ces ressources sont mises en péril par les activités humaines directes comme l’urbanisation et l’agriculture ou indirectes comme le réchauffement climatique.
Références de l’article :