Les jeunes enfants sont dépendants d’une alimentation à base de lait et dans toutes les populations du monde, cela est rendu possible par l’expression du gène codant l’enzyme lactase, chargée de dégrader le lactose en galactose et glucose, des molécules plus simples et facilement incorporables par leur métabolisme. C’est à partir de trois ans que des inégalités apparaissent : certaines populations, dites « lactase persistantes » (LP) restent capables de digérer le lactose, même à l’âge adulte, tandis que d’autres perdent cette capacité du fait que le gène codant la lactase n’est plus exprimé. La répartition des populations LP, fréquentes chez les populations européennes, africaines et arabes, a été largement associée à leurs ancêtres qui ont domestiqué des animaux exploités pour leur lait depuis le Néolithique. L’hypothèse adaptative étant que chez les populations qui disposaient de la ressource en lait, le phénotype LP était avantagé car il apportait un gain nutritionnel, tandis que les individus non LP étaient victimes de crampes intestinales et de diarrhées sévères.

Une jument en cours de traite, dans la vallée de Suusamyr, au Kirghizistan (Crédit : Firespeaker, CC-BY-SA 3.0)
L’article de Laure Segurel et ses collaborateurs français et ouzbeks, publié dans PloS Biology le 8 juin 2020, vise à tester si cette hypothèse largement diffusée est généralisable à tous les peuples d’éleveurs. La comparaison de la fréquence des populations LP et de la carte des populations pastorales, dépendantes de l’élevage, montre une incohérence en Asie centrale (prise au sens large : ex-républiques soviétiques d’Asie centrale mais aussi Mongolie, Chine occidentale et sud-est de la Russie). Les éleveurs kazakhs et mongols ont seulement une fréquence d’individus LP de 12 à 30 %, malgré une consommation importante de produits laitiers. Le génotypage de 30 populations d’Asie centrale montre même que les populations d’éleveurs auraient une fréquence d’individus LP encore plus faible que les populations de fermiers, pourtant moins dépendants du lait dans leur alimentation. Ce résultat vient encore questionner la véracité de l’hypothèse adaptative sur l’ensemble de l’Eurasie.
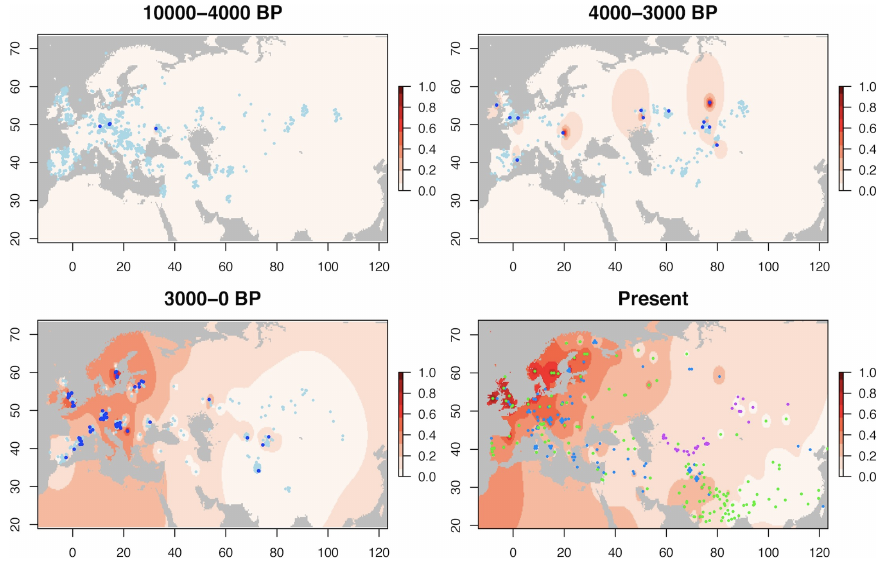
Variation de la fréquence en Eurasie de l’allèle -13.910*T, responsable du phénotype LP, au cours des 10 000 dernières années. La carte de l’Eurasie est indiquée pour quatre périodes : il y a 4 000 à 10 000 ans, il y a 3 000 à 4 000 ans, les 3 000 dernières années (toutes trois alimentées par des échantillons d’Hommes issus de fouilles archéologiques) et enfin l’époque contemporaine (échantillons d’Hommes actuels). Les zones de couleurs représentent une extrapolation de la fréquence de l’allèle -13.910*T : les zones claires représentent des régions avec une faible fréquence de cet allèle (l’autre allèle C y domine) tandis que les zones sombres représentent des régions avec une forte fréquence de cet allèle. Pour les cartes de périodes anciennes, les points bleus foncés représentent l’allèle mutant T tandis que les points bleus clairs représentent l’allèle originel C. On voit sur la première carte que les premiers porteurs de l’allèle muté se situent en Europe centrale et on observe sur les suivantes l’expansion de l’allèle en Eurasie et sa forte augmentation de fréquence en Europe du Nord contrairement à l’Asie centrale où la fréquence allélique reste faible.
Vu que l’adaptation est un processus qui s’inscrit dans la durée, il est important de remettre la problématique dans une perspective temporelle, notamment parce qu’il y a pu y avoir au cours du temps des remplacements de populations. La version du gène responsable du phénotype LP, nommée allèle -13.910*T a ainsi été suivie chez 1 434 individus ayant vécu en Eurasie au cours des 10 000 dernières années. C’est en Europe centrale, il y a 5 950 ans, que l’allèle -13.910*T apparaît pour la première fois. Il se répand ensuite rapidement dans toute l’Eurasie à l’Age de Bronze tardif et on en trouve trace en Asie centrale pour la première fois il y a 3 713 ans. Et c’est à l’Age de Fer que le phénotype LP en Europe et en Asie centrale prend des trajectoires différentes : sur les 3 000 dernières années, la fréquence des individus LP s’élève en moyenne à 31% en Europe alors qu’elle n’est que de 6% en Asie centrale. Cela signifierait donc que l’allèle -13.910*T aurait été fortement sélectionné chez les peuples d’éleveurs d’Europe, notamment l’Europe du Nord, mais pas ou peu sélectionné chez les peuples d’éleveurs d’Asie centrale. Pourquoi ?
Même s’il existe une forte diversité d’animaux domestiques élevés pour leur lait en Asie centrale, le lait de jument occupe depuis très longtemps une place traditionnelle. Paradoxalement, c’est pourtant l’un de ceux dont la teneur en lactose est la plus importante mais c’est aussi l’un de ceux qui est le plus enclin à faire spontanément de la fermentation. Plus généralement, quelle que soit l’origine du lait, les peuples d’Asie centrale ont coutume de ne presque jamais consommer de lait brut, mais plutôt des produits laitiers issus de fermentation, par exemple les boissons fermentées nommées ‘kumis’ ou ‘ajrag’. Dans ces produits fermentés, la teneur en lactose est réduite et, à teneur égale, celui qui reste est plus facilement digéré dans les produits fermentés que dans les produits laitiers non fermentés. D’après l’hypothèse de l’article, si la consommation de produits laitiers en Europe du Nord et en Afrique a pu se faire par une adaptation génétique humaine, l’Asie centrale, où la mutation avantageuse était pourtant présente, a plutôt été marquée par une adaptation culturelle par consommation de produits laitiers fermentés.
Si les peuples d’Asie centrale ne se sont pas eux même adaptés à la consommation de lactose, ils auraient donc exploité les bactéries impliquées dans la fermentation. L’article pose aussi une deuxième hypothèse impliquant encore une fois des bactéries : celle d’un microbiote intestinal qui se serait adapté pour digérer le lactose. La métagénomique, c’est-à-dire l’analyse du contenu en espèces microbiennes dans un environnement donné, par exemple l’intestin, appliquée à des échantillons de différents peuples permettra de tester cette hypothèse.
Finalement, l’une des questions qui reste la plus ouverte est pourquoi y a-t-il pu y avoir adaptation génétique humaine si une autre voie était possible ? Est-ce lié au fait que l’élevage en Europe du Nord et en Afrique est dominé par les espèces bovines (spécificité en terme de quantité/qualité du lait ?) ? Ou est-ce simplement parce que les produits fermentés n’ont pas trouvé d’adeptes dans ces régions ? Nul doute que le généticien des populations ne réussira à répondre à ces questions que dans le cadre de travaux interdisciplinaires et ces passionnants futurs développements seront à surveiller… comme le lait sur le feu.
Références de l’article