Si l’effet de la domestication et de la création variétale sur la diversité des plantes cultivées est largement étudié, l’impact des activités humaines sur les populations de pathogènes affectant les plantes reste beaucoup moins connu. Parmi l’histoire récente de l’agriculture, la Révolution verte est, dans un contexte d’accroissement démographique après la Seconde Guerre mondiale, une période marquée par de profondes transformations du système agricole, parmi lesquelles la croissance exponentielle de la monoculture. Ainsi, aux Philippines, de nombreuses variétés locales ont été cultivées par les fermiers pendant près de 3 500 ans jusqu’à la création dans les années 1960 de l’International Rice Research Institute (IRRI) pour répondre aux enjeux de croissance démographique et de sécurité alimentaire. Les travaux d’amélioration génétique de cet institut ont mené à l’obtention des variétés à haut rendement IR20 en 1967 puis IR64 au milieu des années 1980, toutes deux porteuses du gène Xa4, permettant la résistance à la bactériose du riz, une maladie foliaire causée par la bactérie Xanthomonas oryzae pv. oryzae (appelée ‘Xoo’ dans la suite de l’article). Le succès de ces variétés et de leurs descendantes fut tel qu’elles furent déployées sur des millions d’hectares dans toute l’Asie. Suite à cela, des populations de bactéries Xoo virulentes, c’est-à-dire contournant la résistance, sont apparues aux Philippines.

Symptômes de bactériose sur des feuilles de riz, causés par la bactérie Xanthomonas oryzae pv. oryzae (crédit photo : Donald Groth, publié sur Wikimedia Commons sous licence Creative Commons Attribution 3.0)
Le travail de Ian Lorenzo Quibod et de ses collaborateurs des Philippines, d’Indonésie et du Gabon, publié dans la revue The ISME Journal fin octobre 2019, vise à comprendre les mécanismes de cette apparition de populations virulentes de pathogènes en réponse au déploiement des variétés de riz résistantes. Plus de 70 % des variétés de riz obtenues entre 1960 et 2010 portent la résistance Xa4, alors qu’il existe d’autres sources de résistance qui ont été très peu utilisées (xa5, xa13, Xa7, Xa21). La part de culture des variétés portant le gène de résistance Xa4 a même atteint son maximum en 1988 avec 91,5 % de la surface cultivée en riz aux Philippines portant cette résistance, avant de décroître par la suite, du fait de l’émergence de bactéries Xoo virulentes sur ces cultures. Les tests pathologiques réalisés pour 1822 isolats de bactéries Xoo échantillonnés entre 1970 et 2015 ont montré que la proportion de souches virulentes vis-à-vis des riz portant Xa4 était d’environ 20 % au début des années 1970 puis a rapidement dépassé 50 % jusqu’à la fin des années 1980 avant d’atteindre 84 % au début des années 1990, lors du maximum du déploiement de la résistance Xa4 parmi les variétés de riz cultivées dans l’archipel des Philippines.
L’étude des 11 races de bactéries Xoo identifiées en fonction de leur capacité à se développer sur un panel de variétés de riz portant différentes résistances montre que certaines races sont retrouvées tout au long de l’intensification du déploiement de la résistance Xa4 (races 2 et 3 par exemple) alors que le complexe de races 9com semble, lui, émerger vers le début des années 1990.
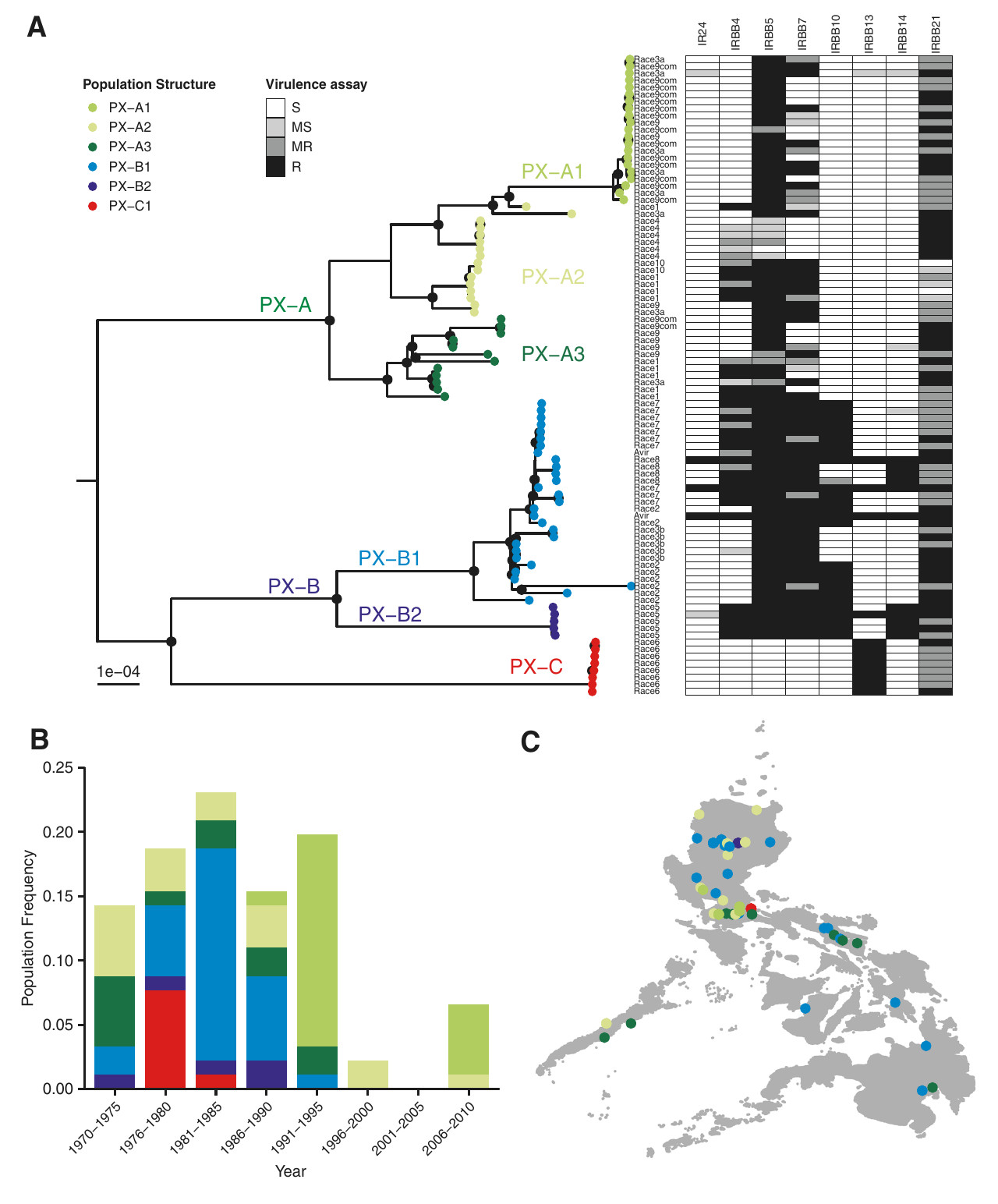
Relations phylogénétiques et représentation temporelle et spatiale des souches de Xanthomonas oryzae pv. oryzae aux Philippines entre 1970 et 2015. A. Arbre phylogénétique construit par comparaison des régions génomiques communes entre souches. Selon la topologie de l’arbre, chaque souche est ainsi classée dans l’un des six groupes génétiques identifiés : PX-A1, PX-A2, PX-A3, PX-B1, PX-B2, PX-C1. En plus d’être comparées au niveau génomique, les souches Xoo ont aussi été comparées en terme de virulence vis-à-vis de huit variétés de riz présentant chacune un gène de résistance Xa différent (en haut à droite de la figure). Chaque variété de riz est ainsi indiquée comme sensible (S), moyennement sensible (MS), moyennement résistante (MR) ou résistante (R) à la souche correspondante. En fonction de leur capacité à infecter ces variétés de riz témoins, les souches sont classées en races. B. Distribution dans le temps de la proportion de souches de bactéries Xoo isolées entre 1970 et 2015. Les souches sont indiquées selon le groupe génétique qui leur a été assigné. Les couleurs du graphique sont les mêmes que celles de l’arbre phylogénétique. Les périodes de temps réunissent les souches échantillonnées sur cinq années consécutives. L’axe des ordonnées indique la proportion sur l’ensemble des 91 souches étudiées sur la période 1970-2015. On notera par exemple que les souches, non virulentes, du groupe PX-B2 (violet) ont fini par disparaître au cours du suivi, tandis que les souches, virulentes, du groupe PX-A1 (vert le plus foncé), absentes au début du suivi, ont émergé au cours de l’étude, en parallèle du déploiement de la résistance. C. Distribution spatiale des souches de Xoo dans l’archipel des Philippines. Les souches sont représentées en fonction de leur assignation aux groupes génétiques. Les couleurs sont les mêmes que pour l’arbre phylogénétique.
Afin de comprendre les changements génomiques des bactérie sur cette période, le génome de 91 souches de Xoo a été séquencé. La comparaison de ces génomes a permis de distinguer six groupes génétiques : PX-A1, PX-A2, PX-A3, PX-B1, PX-B2 et PX-C1. Ces groupes étaient très variables en terme de mutations et de recombinaisons, montrant une histoire évolutive complexe. Plusieurs variations nucléotidiques étaient associées à la virulence vis-à-vis du gène Xa4, notamment pour des gènes impliqués dans les systèmes de sécrétion, la dégradation de la paroi cellulaire, la détoxification des Reactive Oxygen Species (ROS) et la production de lypopolysaccharides. Plusieurs de ces variations se sont aussi avérées être sous sélection purifiante, c’est-à-dire comme subissant une intense contre-sélection contre les mutations affectant la fonction des gènes correspondants. La multiplicité des gènes candidats associés à l’émergence de la virulence vis-à-vis de la résistance Xa4 suggère que plusieurs acteurs moléculaires différents auraient pu évoluer en réponse au déploiement de la résistance Xa4 parmi les variétés de riz cultivées.
Globalement, l’étude montre plusieurs destinées possibles des souches de bactéries Xoo avant, pendant et après le déploiement des variétés de riz portant la résistance Xa4. Les souches de Xoo non virulentes, par exemple celles du groupe génétique PX-B2, en forte fréquence avant le déploiement, ont fini par disparaître du fait qu’elles n’étaient plus capables d’infecter les nouvelles variétés de riz résistantes. Les souches virulentes vis-à-vis de la résistance Xa4, comme celles du groupe génétique PX-B1, notamment la race 2, déjà présentes en fréquence limitée avant le déploiement, car non avantagées, ont par la suite été avantagées par le déploiement de la résistance Xa4 et ont augmenté en fréquence. Enfin, des souches inconnues avant le déploiement, comme celles du groupe génétique PX-A1, notamment le complexe de races 9com, ont pu émerger suite au déploiement.
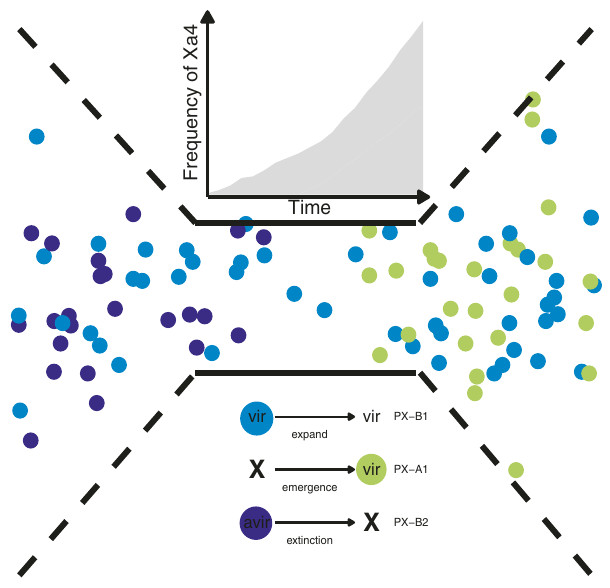
Schéma synthétique montrant l’effet du déploiement progressif au cours de la Révolution verte des variétés de riz portant la résistance Xa4 (cf. graphique) sur l’évolution des fréquences de souches de Xanthomonas oryzae pv. oryzae. Avant la Révolution verte (gauche du schéma), les souches virulentes (bleu) et les souches non virulentes (violet) co-existaient. Puis le déploiement de la résistance chez le riz a agi comme un goulot d’étranglement sur la diversité des souches pré-existantes : les souches non virulentes, incapables d’infecter ces nouvelles variétés de riz résistantes, ont disparu tandis que les souches virulentes ont augmenté en fréquence car elles étaient les seules à pouvoir se développer dans les cultures de riz. Cette forte pression de sélection sur les pathogènes a même pu entraîner l’apparition de nouvelles souches virulentes (vert), jusqu’alors inconnues dans l’archipel des Philippines. A droite, le goulot d’étranglement a cessé quand la résistance Xa4 n’a plus été systématiquement incorporée aux nouvelles variétés de riz cultivées dans l’archipel. Toutefois, la composition des populations de pathogènes avait drastiquement changé entre avant et après le goulot d’étranglement.
La prise en compte de la co-évolution entre la plante hôte et ses pathogènes dans un contexte de changement de pratiques agricoles est une question primordiale pour la sécurité alimentaire. Des études comme celle-ci permettront peut-être de mieux réfléchir aux stratégies futures de déploiement spatial et temporel des sources génétiques de résistance, à l’avenir.
Références de l’article