
Chez les espèces à multiplication sexuée, seules les mutations touchant les cellules germinales sont transmissibles à la descendance. Selon un modèle sélectionniste, ce postulat doit conduire à observer des taux de mutations variables selon les lignées cellulaires. Ainsi, chez les animaux, les cellules germinales, dont la lignée apparaît tôt au cours du développement, montrent un taux de mutation plus réduit que les cellules somatiques. Le taux de mutation est aussi inversement proportionnel à la longévité des lignées de cellules somatiques : les lignées de cellules somatiques des animaux à faible longévité montrent ainsi un taux de mutation plus élevé que celles des animaux à forte longévité.
L’article de Wang et al. 2019 paru dans la revue PLoS Biology tente d’explorer la généralisation de ces conclusions aux végétaux et d’en identifier les spécificités. Pour ce faire, 754 génomes ont été séquencés, correspondant à différents tissus étudiés chez huit espèces de plantes : parmi les plantes pérennes, les pêchers cultivés (Prunus persica) et sauvage (Prunus mira), l’abricotier du Japon (Prunus mume), le fraisier des bois (Fragaria vesca) et une espèce de saule (Salix suchowensis), tandis que pour les plantes annuelles, du riz cultivé (Oryza sativa), la graminée Brachypodium distachyon et l’arabette des dames (Arabidopsis thaliana).
Un des principaux facteurs qui influencent le taux de mutation est la transmissibilité des mutations. Contrairement aux animaux, il y a encore beaucoup de débat pour savoir si les plantes disposent d’une lignée cellulaire germinale clairement et durablement indépendante des lignées somatiques, apparaissant tôt dans le développement. Toutefois, on peut faire la distinction entre les cellules des tiges et celles des racines, puisque seules les premières pourront contribuer à des lignées germinales, à travers la formation de bourgeons floraux. L’article montre ainsi que les tiges ont un taux de mutation plus faible que les racines chez les espèces pérennes. La différence n’est en revanche pas significative chez les plantes annuelles, pour lesquelles les mutations survenant sur les tiges ne disposent probablement pas des mêmes probabilités de transmission à la descendance. Chez les plantes annuelles, pour lesquelles les mutations pré-méiotiques (apparues dans des cellules somatiques des tiges) ont peu de chances d’être transmises, il n’y a pas possibilité d’accumulation de génération en génération et par conséquent il n’y a pas de nécessité sélective à réduire le taux de mutation dans les tiges chez les plantes annuelles.
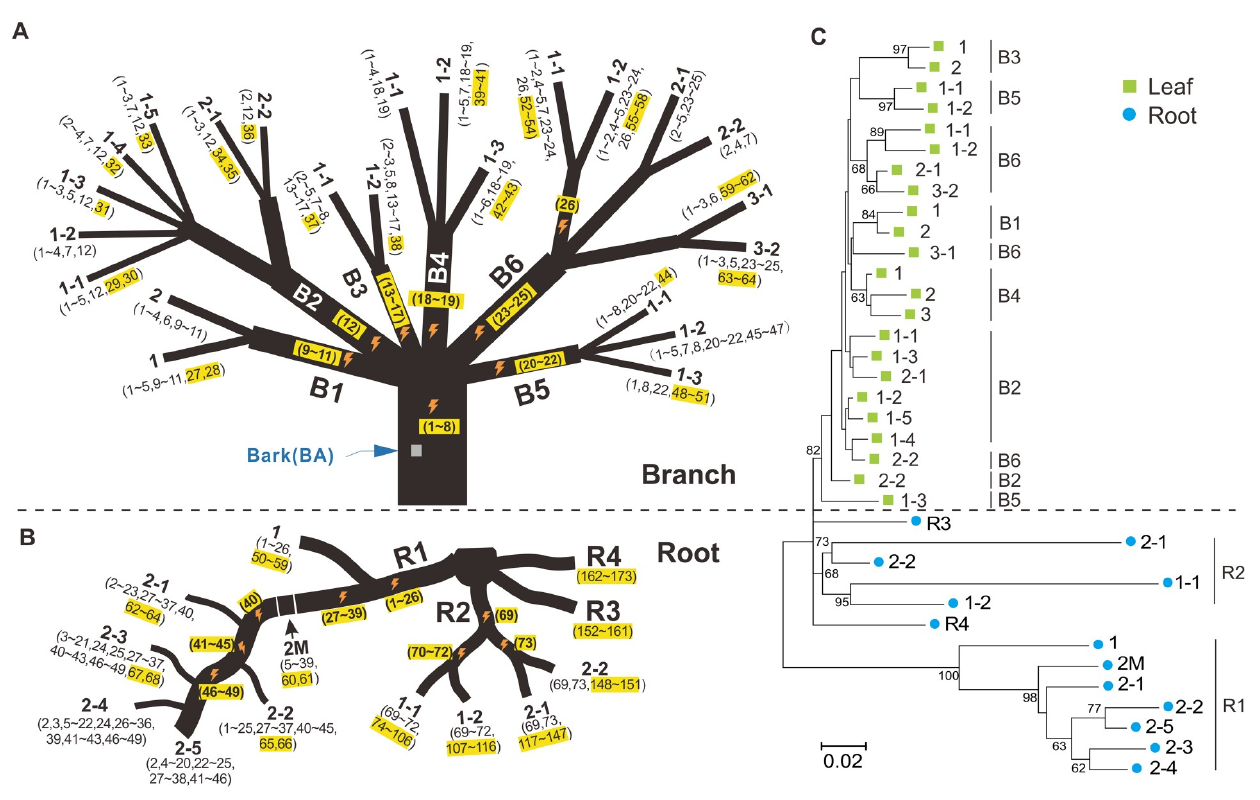
Événements de mutation détectés dans les feuilles et les racines d’un pêcher. (A) Mutations somatiques des feuilles. Les mutations sont numérotées et entre parenthèses. La première présence ontogénétique d’une mutation est surlignée en jaune. Les éclairs symbolisent la présence de mutations survenues dans des branches internes. (B) Mutations somatiques des racine, représentées de la même façon. (C) Arbres ontogénétiques construit à partir de toutes les mutations identifiées dans les branches (vert) ou les racines (bleu). Comme pour un arbre phylogénétique, les longueurs de branche sont proportionnelles à la distance génétique. On voit ici nettement que les racines sont plus divergentes génétiquement entre elles que ne sont les feuilles.
Les auteurs ont identifié une exception à cette construction intellectuelle : les stolons de fraisiers. Les stolons sont des tiges rampantes à partir desquelles des bourgeons latéraux peuvent initier de nouvelles plantes ayant leurs propres racines. L’étude montre que ces stolons de fraisiers ont un taux de mutations anormalement élevé pour une tige de plante pérenne, selon la logique sélectionniste expliquée plus haut. Mais il semblerait que la probabilité qu’une mutation survenant dans le stolon soit transmise aux bourgeons latéraux et ainsi aux plantes filles, soit extrêmement faible. Ce résultat laisse suggérer que les lignées cellulaires des stolons et celles des bourgeons latéraux seraient ontogénétiquement indépendantes.
Les auteurs ont aussi essayé d’écarter l’hypothèse que les différences de nombre de mutations observées ne soient pas expliquées par des variations de taux de mutations, mais plutôt, à taux de mutations constant, par des variations de l’intensité de la sélection purifiante qui éliminerait les mutations les plus désavantageuses pour le devenir de la plante. Deux raisons ont mené à écarter cette hypothèse : (i) les mutations somatiques s’accumulent à un même taux quel que soit l’âge du tissu alors que la sélection purifiante, qui est un processus qui se passe sur le long terme, devrait être plus visible sur les tissus anciens, et (ii) il n’y a pas de démonstration que les mutations potentiellement plus délétères (ex : mutations non synonymes, c’est-à-dire qui modifient la séquence en acides aminés) soient moins conservées.
Les auteurs concluent donc que ce serait avant tout directement le taux de mutations qui serait sous sélection, et pas les mutations générées. Parmi les autres facteurs qui viennent appuyer ce mécanisme, on trouve la longévité des lignées cellulaires : ainsi des lignées cellulaires éphémères comme les pétales ont un taux de mutations plus élevé par comparaison à des lignées cellulaires plus durables comme les feuilles, probablement pour limiter les effets d’accumulation de mutations chez ces organes plus durables. Cependant, on doit garder en tête que des facteurs environnementaux peuvent aussi affecter ce taux de mutations. Un taux très élevé de mutations a ainsi été observé chez des plantes issues de culture in vitro, confirmant qu’un stress pouvait anormalement augmenter le taux de mutations dans les cellules.
Ce travail est une illustration du concept de métapopulation à l’intérieur d’une plante : le génome varie à l’intérieur d’une plante, du fait des mutations accumulées lors de son développement. Parmi les enjeux appliqués de ces travaux, on peut penser à la compréhension des mutants végétatifs ou sports, c’est-à-dire de branches de phénotype différent des autres branches d’un individu et qu’on peut potentiellement propager par la suite de manière végétative (bouturage, greffage). Elles ont joué un rôle important dans la création variétale de nombreuses espèces pérennes cultivées comme la vigne, le pommier ou le rosier.

Cultivar nain d’épinette blanche (Picea glauca var. albertiana ‘Conica’) dont une branche a réverté vers le phénotype initial.
Source : Ragesoss, Wikimedia sous licence Creative Commons
Références de l’article :