
En 1956, le généticien Haldane posait la question suivante à des anthropologues : « Les différences biologiques entre les groupes humains sont-elles comparables avec celles de groupes d’animaux domestiques, tels que les lévriers ou les bulldogs ?». Les prolongements de cette question ont donné lieu à un débat populaire qui connaît son paroxysme dans des décisions politiques, qui sans revenir à l’idéologie nazie, ont abouti aux cinq catégories raciales actuellement en vigueur dans la législation américaine (Blanc /Noir ou Afro-Américain / Amérindien ou natifs d’Alaska /Asiatique / Hawaïen ou natifs d’autres îles du Pacifique). Mais le concept de races humaines ne serait-il pas davantage une construction sociale plutôt qu’une réalité biologique comparable aux races d’animaux domestiques, notamment les races canines ?
A gauche, planche de races de chiens, tirée de ‘The New Student’s Reference Work’ (Ed. Chandler B. Beach, Chicago) publié en 1914. A droite, planche de visages humains d’origine variée, tirée de ‘Evolution of Life’ par Henri C. Chapman en 1873 (Ed. J.B. Lippincott, Philadelphie). Illustrations du Domaine public.
C’est la question à laquelle la chercheuse américaine Heather L. Norton et ses collaborateurs ont tenté de répondre dans un article de synthèse paru le 9 juillet 2019 dans la revue Evolution: Education and Outreach. Le concept génétique de races repose sur le fait qu’il existerait des groupes distincts au sein d’une espèce, c’est-à-dire pour lesquels la diversité intra-groupe serait minime alors que la diversité inter-groupes serait très importante. Chez l’Homme, les anthropologues ont comparé les individus par rapport à des critères comme la couleur de la peau, des mesures de crânes, les groupes sanguins ou encore des marqueurs génétiques.
Ainsi, l’analyse de la diversité génétique de chiens à partir de marqueurs moléculaires classait les individus dans des groupes génétiques qui correspondaient aux races dans 99 % des cas. Le même type d’analyse mené sur un groupe d’humains échantillonnés sur l’ensemble de la planète aboutissait au modèle le plus vraisemblable distinguant six groupes génétiques répartis en : (1) Afrique, (2) Europe/Moyen-Orient/Asie centrale, (3) Extrême-Orient, (4) Océanie, (5) Amériques et (6) les Kalashs, une population isolée du Nord-Ouest du Pakistan. La plupart de ces groupes génétiques s’explique par la barrière reproductive que représentent les limites entre continents. Toutefois, contrairement aux chiens, ces groupes génétiques ne doivent pas être interprétés comme des races. Déjà, l’analyse n’aboutit pas à un modèle incontestable de structure et il existe plusieurs modèles plausibles. On peut même parler de modèles imbriqués, indiquant plusieurs niveaux de structuration génétique plus ou moins marqués. Ainsi, si on sépare l’humanité en deux groupes on distinguera les populations d’Afrique/Europe/Moyen-Orient/Asie centrale de celles de l’Extrême-Orient/Océanie/Amériques. Si on sépare l’humanité en trois groupes, les populations d’Afrique de distinguent des populations d’Europe/Moyen-Orient/Asie centrale. Le découpage en quatre groupes génétiques sépare les populations d’Amériques de celles d’Extrême-Orient/Océanie. Ce dernier groupe est séparé dans le cas d’une subdivision en cinq groupes génétiques. Par ailleurs, peu importe le découpage, la probabilité d’assignation d’un humain à un groupe génétique n’est souvent pas totale et une personne est généralement assignée à plusieurs groupes génétiques simultanément.
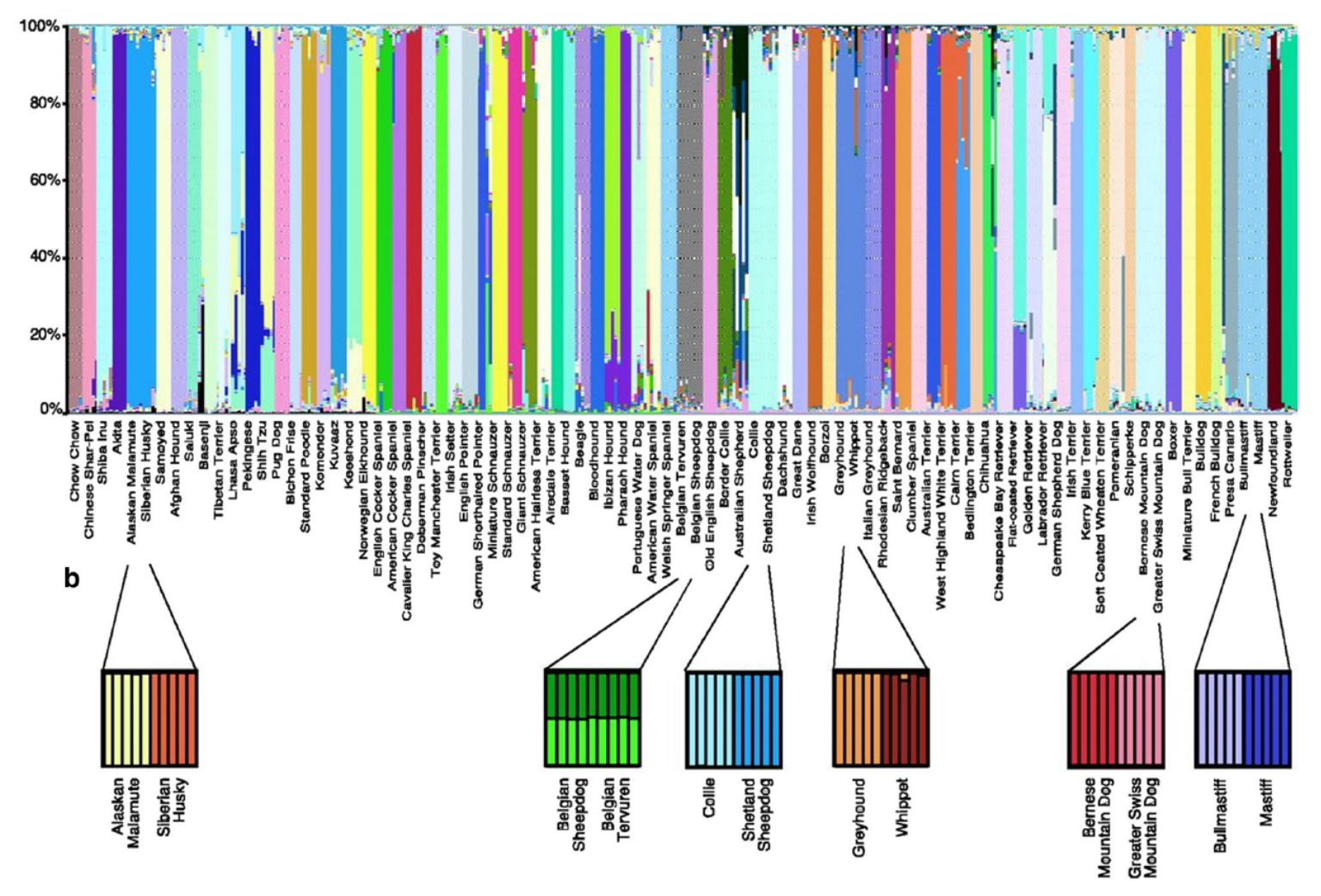
Structure génétique observée pour 85 races de chiens. (a) L’abscisse montre les 85 races de chiens représentées par plusieurs individus. L’ordonnée indique la probabilité d’assignation aux groupes génétiques, représentés par des couleurs différentes. Les groupes génétiques identifiés sont généralement cohérents avec les races canines. (b) Analyses effectués indépendamment sur des couples de races que l’analyse globale n’avait pas réussi à différencier car ces races ont une origine commune (ex : Mastiff et Bullmastiff). A l’exception de deux types de Bergers Belges (le Groenendael et le Tervueren), ces analyses sur des sous-échantillons ont permis de différencier génétiquement les races. Cette figure a initialement été publiée dans Parker et al. (2004).
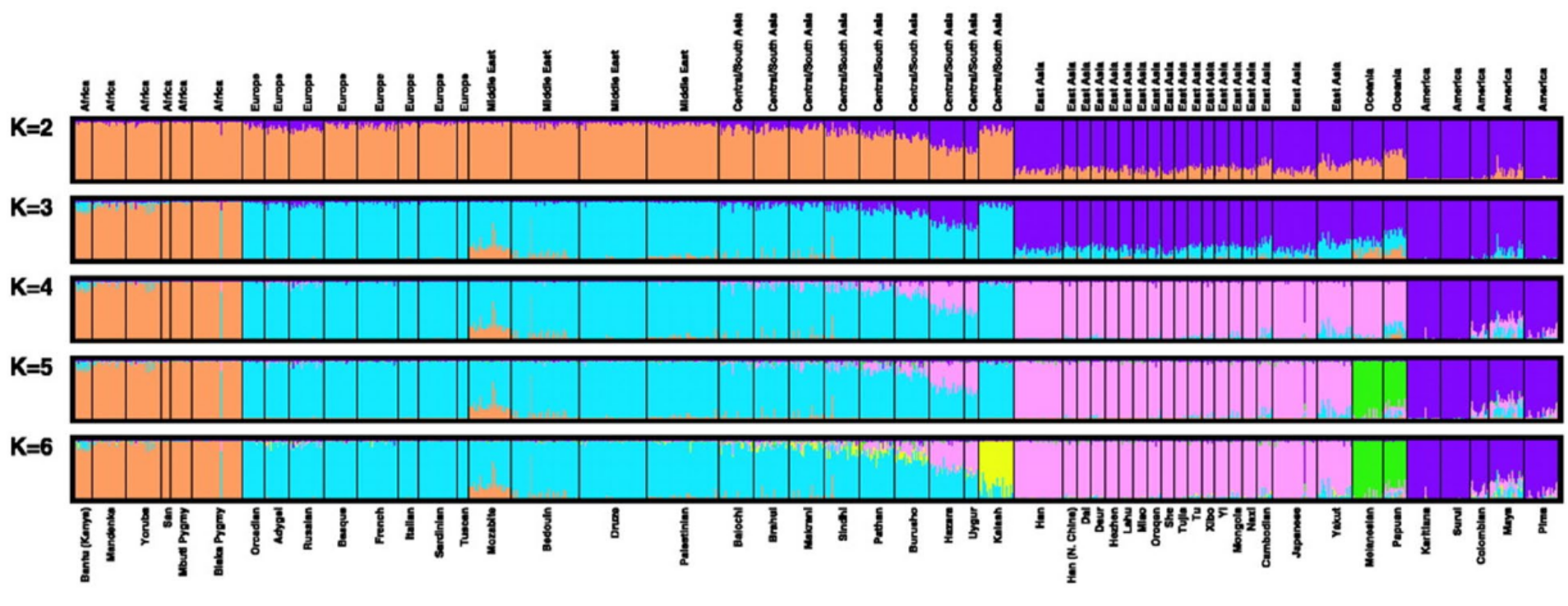
Structure génétique observée pour 52 populations humaines échantillonnées dans le monde entier. Les cinq analyses effectuées en fonction du nombre de groupes génétiques (K=2 à 6) montrent ce concept de structure imbriquée. L’origine géographique, notamment l’appartenance à un continent, est le facteur le plus structurant. Toutefois, de nombreux individus sont assignés à plus d’un groupe génétique. Ces résultats ont été publiés pour la première fois dans un article de Rosenberg et al. 2002.
L’analyse de la variance moléculaire (AMOVA) est un outil statistique permettant de décomposer la variance génétique selon différents niveaux hiérarchiques de structuration. Elle s’avère donc un bon moyen de tester la pertinence des groupes génétiques détectés. Si 27 % des différences génétiques observées entre les chiens se situent entre races canines, seulement 3,3 à 4,7 % de la variation génétique totale observée chez les humains se situent entre les groupes continentaux ou régionaux évoqués plus haut.
Comment expliquer ces différences entre la structuration des populations canines et des populations humaines ? La plupart des races de chiens ont une origine récente, correspondant à environ cent ans de sélection drastique où l’Homme a empêché les chiens de se reproduire en dehors de la race et n’a autorisé les reproductions qu’entre un petit nombre d’individus correspondant aux standards fixés pour la race. Il en résulte une diversité réduite qu’on peut estimer par l’hétérozygotie attendue H, c’est-à-dire la probabilité de tirer deux allèles différents connaissant les fréquences alléliques de la population. Chez une race canine, H est compris entre 0,313 et 0,610. Cette consanguinité est par ailleurs à l’origine d’un nombre considérable de maladies génétiques propres à chaque race. Du côté de l’espèce humaine, les facteurs qui empêchent les flux de gènes entre groupes régionaux sont des facteurs géographiques, culturels et linguistiques, qui, même s’ils s’exercent depuis plus longtemps que la sélection des races modernes de chiens, n’a pas eu un effet d’isolement génétique aussi marqué. Il en résulte une diversité génétique intra-groupe plus élevée que celles des races canines (H=0,664-0,792). L’indice de fixation FST qui marque l’ampleur de la différentiation génétique entre des populations était aussi largement plus élevé entre races canines (FST=0,33) qu’entre groupes continentaux humains (FST=0,052-0,083).
Le concept de race suppose aussi une homogénéité phénotypique. Un caractère aussi stigmatisant que celui de la couleur de la peau chez l’humain varie en fait assez largement au sein d’un continent puisque les populations originaires des tropiques et des hautes altitudes présentent les couleurs de peau les plus sombres, afin d’apporter une forte protection contre les UV. Ces continuums de couleurs de peau sont aussi permis par un contrôle génétique complexe de ce caractère de couleur de peau chez l’Homme (des centaines de gènes impliqués) contrairement aux chiens (neuf gènes identifiés). Quant à la taille des individus, leurs distributions se chevauchent largement entre catégories raciales américaines. Cela contraste fortement aux très impressionnantes différences de taille entre les races de chiens.
Alors, que reste-t-il au concept de race chez l’espèce humaine ? Une construction sociale bâtie en parallèle du racisme permettant à une élite dominante de stigmatiser des minorités. Preuve en est que ces classes raciales ont évolué au cours du temps pour appuyer des motivations politiques telles que l’esclavage ou les politiques migratoires. On peut définir le racisme comme l’idéologie politique selon laquelle les groupes humains ne seraient pas dotés des même capacités et qu’on leur attribuerait un jugement de valeur en distinguant des races supérieures et des races inférieures. L’historien Harrington considère que l’apparition du concept de races humaines aux États-Unis n’est pas étrangère à la généralisation dans les années 1880 des races pures de chiens, par opposition à ce qu’on appelle aujourd’hui les ‘village dogs’ qui sont aux chiens ce que le ‘chat de gouttière’ est aux chats. Cet événement coïncide avec le rejet des immigrants irlandais, allemands, italiens, juifs par les White Anglo-Saxon Protestants (WASP) déjà présents aux États-Unis.
Références de l’article :